Abstract
Background: Discoid lupus erythematosus is a cutaneous disease with a worldwide distribution, and its pathogenesis remains unclear. Case Report: A 41 year old male was evaluated for hair loss, in patches on the scalp. We studied selected adaptor proteins expressed in T, natural killer, neutrophil and mast cells; these proteins are important mediators for antigen receptor signaling in situ. Methods: Skin biopsies for hematoxylin and eosin examination, as well as for direct immunofluorescence and immunohistochemistry analysis were performed. Results: Hematoxylin and eosin staining demonstrated classic features of lupus with focal dermal scarring; epidermal atrophy was noted, with lymphohistiocytic infiltrates around the skin appendages. Direct immunofluoresence revealed classic, lupus band positive staining along the dermal/epidermal junction. In addition, immune reactants were identified in neurovascular areas, and around pilosebaceous units. Immunohistochemistry staining showed positive staining for the T-cell antigen receptor zeta chain, the linker for activation of T cells, myeloperoxidase, cyclo-oxygenase 2, melanoma-associated antigen 1, B cell leukemia/lymphoma-2 associated X protein, and BCL-2 markers. The positive staining was observed within the dermal inflammatory infiltrate, around pilosebaceous units, upper dermal blood vessels, and focally within eccrine sweat glands. Conclusions: The pathobiology of cutaneous lupus involves not only the epidermis, but also dermal pilosebaceous units, eccrine sweat glands and blood vessels. Further studies are recommended, especially in the light of presented data regarding T cell activation and proapototic molecules.
Key words: cutaneous lupus; ZAP-70; LAT; BCL-2; myeloperoxidase; COX-2; MUM-1; pilosebaceous unit; sweat glands
Introduction
Cutaneous lupus erythematosus (LE) covers a broad morphological spectrum, extending beyond acute, subacute and chronic cutaneous lupus erythematosus, which are commonly classified as lupus-specific skin disease [1]. Other, less common presentrations include tumid lupus erythematosus, lupus profundus, chilblain lupus, mucosal lupus erythematosus and bullous lupus erythematosus [1]. Possible vascular sequelae of lupus erythematosus include leukocytoclasis, urticarial vasculitis, livedoid vasculopathy and livedo reticularis [1]. Many previous histologic investigations have emphasized 1) immune deposits at the dermoepidermal junction (DEJ) of the base membrane zone (BMZ), i.e., the lupus band), and 2) the role of autoantibodies and antibody dependent cellular cytotoxicity in the pathogenesis of LE [1]. It is known that autoimmune T helper cells drive pathogenic autoantibody production in LE, but the mechanisms maintaining those pathogenic T cells are unknown. Here, we explore pertinent T cell signaling activators, and the possible roles of other immune cells mediators in lupus erythematosus. We include products derived from activated neutrophils, such myeloperoxidase; and also other molecules located in a lupus susceptibility region on chromosome 1, such as cyclo-oxygenase 2.
Case Report
A 41 year old male was evaluated for the presence of hair loss, in patches in the scalp. On physical exam, the patient demonstrated atrophic scaly scalp plaques, with hair loss inside the plaques. A lesional skin biopsy was taken for hematoxylin and eosin (H&E) analysis. In addition, direct immunofluorescence (DIF) and immunohistochemistry (IHC) studies were performed.
Methods
In brief, skin preparations for H&E, DIF and IHC studies were performed as previously described. We utilized antibodies from Dako (Carpinteria, California USA), including 1) anti-human cyclo-oxygenase 2 antibody (COX-2), which does not crossreact with cyclooxygenase 1, 2) the T-cell antigen receptor zeta chain (ZAP-70 antibody), 3) the linker for activation of T cells (LAT), myeloperoxidase, 4) mutated melanomaassociated antigen 1 (MUM-1), 5) B cell leukemia/lymphoma-2 associated X protein (BAX) and 6) BCL-2 antibody. The direct immunofluorescence (DIF) and immunohistochemistry (IHC) studies performed as previously described [2-10].
Results
Microscopic description:
Examination of the H&E tissue sections demonstrated classic features of lupus erythematosus; focal atrophy and follicular plugging were noted within the epidermis. The presence of a mild, perifollicular concentric fibrosis and in few areas with some scarring was seen. No significant interface inflammation was noted. A mild, superficial and deep, perivascular and periadnexal dermal infiltrate of lymphocytes, lymphocytes, plasmacytoid lymphocytes and histiocytes was noted. Neutrophils and eosinophils were rare. Mild, perifollicular concentric fibrosis was observed, with additional areas of interstitial scarring. The Verhoeff elastin special stain confirmed the extent of dermal scarring (Fig. 1,2). Focal edema was also appreciated around sebaceous and eccrine sweat glands. DIF demonstrated the following staining results: IgG (++, focal and linear at the epidermal basement membrane zone (BMZ), sebaceous gland BMZs and within selected neurovascular structures; IgA (-); IgM (+, focal, linear epidermal BMZ and dermal perivascular); IgE (+, in selected papillary dermal cells; Complement/C1q (++, focal and linear at epidermal BMZ and sebaceous gland BMZs); Complement/C3 (++, focal linear epidermal BMZ and sebaceous gland BMZs, and surrounding some dermal neurovascular structures; Complement/C4 (++, focal perifollicular and surrounding dermal neurovascular structures); Kappa and Lambda light chains (+, focal punctate epidermal BMZ, and surrounding dermal neurovascular structures and sweat glands); Albumin (++, focal linear epidermal BMZ and sebaceous gland BMZs); fibrinogen (++, focal linear epidermal BMZ and sebaceous gland BMZs) and, finally, ZAP-70, LAT, myeloperoxidase, COX-2, MUM- 1, and BCL-2 (+), around dermal sebaceous and sweat glands, and dermal blood vessels (Fig. 1,2). In addition, some of these final antibodies clearly reacted with the BMZs of the sebaceous and sweat glands.
Discussion
Consistent histopathologic features in many cutaneous lesions of LE include a perivascular mononuclear cell infiltrate, with subsequent involvement of the epidermis and dermal appendages. The histologic alterations affecting the epidermis, dermis, and adnexal structures reflect the specific lesion biopsied, and lesional age at the time of biopsy [11,12]. The multiple clinical and serologic forms of lupus erythematosus cannot reliably be distinguished histologically, which supports the premise that lupus erythematosus is a disease presenting a wide spectrum of clinical manifestations and a common underlying pathogenesis [11,1 2]. In our case, we were able to observe how skin adnexal structures and dermal blood vessels interfaced with the pathologic inflammatory infiltrate; specifically, T cells and neutrophil activated, downstream cell signaling molecules seem to play significant roles in the pathophysiology. Most of the markers of immune activation were identified in the adnexal structures. We were able to identify the presence of COX-2, an inducible enzyme that is normally absent in skin cells; however, in response to growth factors, tumor promoters and some cytokines, it exhibits a rapid and transient expression. In addition, ZAP-70 plays a role in lymphocyte activation. It is known that autoimmune T helper cells drive pathogenic autoantibody production in systemic lupus erythematosus (SLE); however, the pathologic mechanisms maintaining these T cells are unknown. Here we demonstrated both TCR (T-cell antigen receptor) mediated and pre-TCR mediated signaling, both in developing and mature T lymphocytes. LAT is involved in FcGR3 (low affinity immunoglobulin gamma Fc region receptor III)-mediated signaling in natural killer cells, as well as FcER1 (high affinity immunoglobulin epsilon receptor)-mediated signaling in mast cells. Coupled activation of these receptors and their associated kinases would then result in downstream intracellular events, including mobilization of intracellular calcium stores, PKC activation, MAPK activation and/or cytoskeletal reorganization through the recruitment of PLCG1, GRB2, GRAP2, and other signaling molecules.
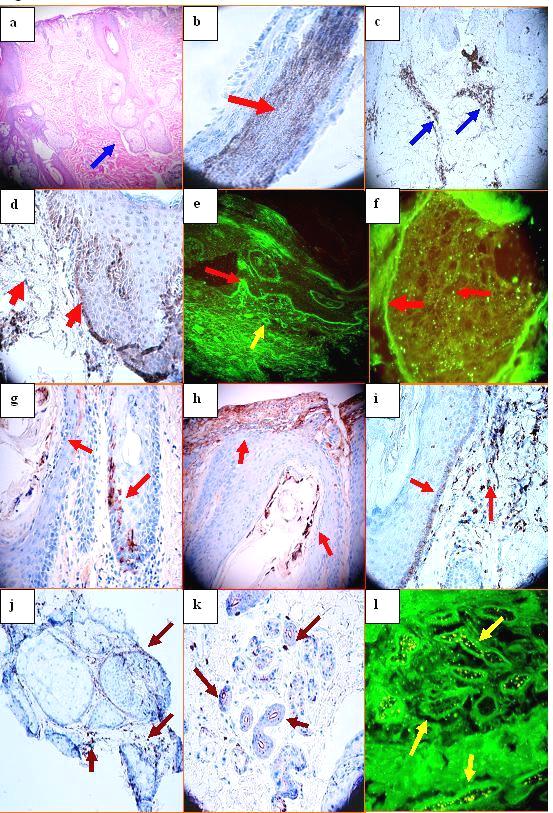 Figure 1. a. H&E, showing atrophy of the epidermis, follicular plugging and no significant hyperkeratosis Mild, perifollicular concentric fibrosis is present, with additional focal interstitial scarring. A mild, superficial, perivascular and periadnexal dermal inflammatory infiltrate is also noted. Focal edema is present within the dermis, especially around hair follicular units (blue arrow). b. Positive staining within a hair follicle with the MUM-1 antibody (brown staining; red arrow). c. Positive staining with LAT antibody on an upper dermal perivascular infiltrate (brown staining; blue arrows). d. Positive staining with anti-BAX antibody accentuated at the basement membrane zone (BMZ) of the epidermis, as well as on individual cells within the papillary dermis(brown staining; red arrows). e. Positive staining with FITC conjugated anti-human-Complement/C3 against the BMZ of the epidermis (green staining; red arrow) and against upper dermal blood vessels (green staining; yellow arrow). f. Positive staining with FITC conjugated anti-human-complement/C3 against the BMZ of a sebaceous glands and some areas within the gland (green staining; red arrows). g. IHC positive staining of a hair follicle with complement/C3, at the BMZ of the hair follicle and inside the hair follicle (brown staining; red arrows). h. Complement/C1q positive IHC staining inside the hair follicle shaft and at the epidermal BMZ area (brown staining; red arrows). i, COX-2 positive staining at the BMZ of a hair follicular unit and in a perifollicular inflammatory infiltrate(dark staining; red arrows). j. IHC positive staining with BCL-2 on inflammatory cells around sebaceous glands (dark staining; maroon arrows). k. IHC positive staining with ZAP-70 inside an eccrine gland coil (dark staining; maroon arrows). l. DIF positive staining against eccrine sweat glands using FITC conjugated anti-human IgG (green staining; yellow arrows)
|
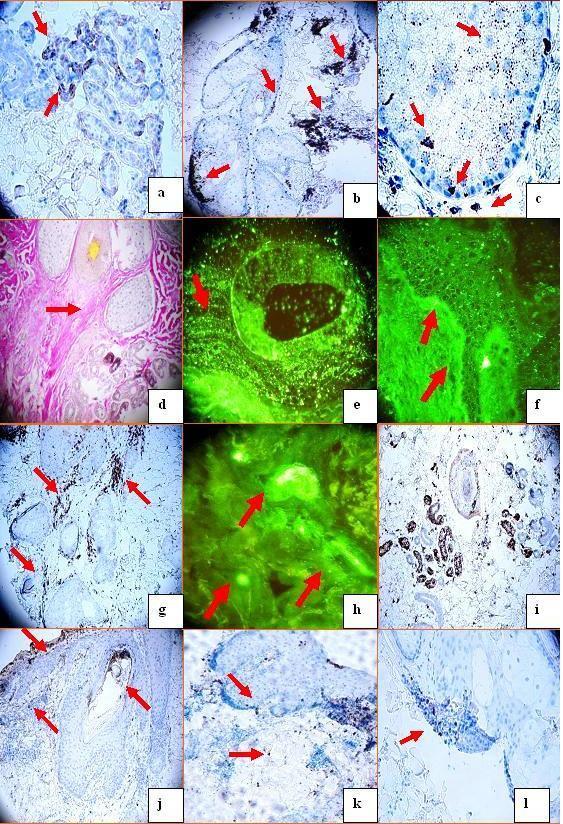 Figure 2 a. Positive IHC staining using anti-human BCL-2 antibody against some areas of an eccrine sweat glands (darkbrown staining; red arrows), b. Positive anti-human-ZAP-70 antibody staining on a sebaceous gland. Please notice how some staining areas are accentuated along the BMZ of the gland, and some staining areas accentuated within the gland interior(brown staining; red arrows). c. ZAP-70 positive IHC staining in multiple patterns within and around a sebaceous gland. The patterns include dots inside and outside the gland, clustered dot staining, and along the gland BMZ (brown staining; red arrows). d. A Verhoeff elastin special stain confirms the extent of dermal scarring (red arrow). e, DIF positive staining with FITC conjugated anti-human-IgG, in a punctate dot pattern around a hair follicle unit (yellow staining; red arrow). f, Positive DIF lupus band staining at the epidermal BMZ with FITC conjugated anti-human-complement/C3( faint yellow staining; red arrows). g Positive IHC staining with anti-human LAT antibody around a hair follicular unit (brown stain, red arrows). h. Positive DIF staining with FITC conjugated anti-human-Complement/C3 against some neurovascular structures (yellow/green staining; red arrows). i. Positive IHC staining with anti-human COX-2 in a hair follicle and the eccrine gland coils (brown staining). j. Positive follicular and perifollicular IHC staining with anti-human myeloperoxidase (brown staining; red arrows). k. Positive IHC staining with anti-human myeloid histiod antigen antibody within and near a sebaceous gland (brown staining; red arrows). l. Positive IHC staining with anti-human MUM-1 antibody around a hair follicle (brown staining; red arrow).
|
Autoreactive T cells are normally eliminated by 1) functional inactivation (anergy) and 2) activationinduced cell death (AICD; directed apoptosis) through death receptor (Fas) signaling. Other authors have reported that activated T cells of lupus patients resist anergy and apoptosis by markedly upregulating and sustaining COX-2 expression [13]. Inhibition of COX-2 caused apoptosis of the anergy-resistant lupus T cells by augmenting Fas signaling and markedly decreasing the survival molecule c-FLIP (cellular homolog of viral FLICE inhibitory protein) [13]. Studies with COX-2 inhibitors and COX-2 deficient mice confirmed that this COX-2/FLIP antiapoptosis program has been used selectively by anergy-resistant lupus T cells, and not by cancer cells or other autoimmune T cells [13]. Notably, the gene encoding COX-2 is located in a lupussusceptibility region on chromosome 1. The same authors also found that selected COX-2 inhibitors were able to suppress the production of pathogenic autoantibodies to DNA by causing autoimmune T-cell apoptosis, an effect that was independent of prostaglandin E2 (PGE2) [13]. These findings could be useful in the design of lupus therapies. Our COX-2 data is consistent with expression of this molecule in the context of clinical lupus erythematosus. The ZAP-70 gene encodes an enzyme belonging to the tyrosine kinase protein family; ZAP-70 plays roles in both T lymphocyte development and activation [14]. The ZAP-70 enzyme, which is phosphorylated on its tyrosine residues upon T cell antigen receptor (TCR) stimulation, functions in the initial step of TCR-mediated signal transduction in combination with Src family kinases. Mutations in the ZAP-70 gene result in a form of severe combined immunodeficiency (SCID) syndrome in humans [14]. ZAP-70 expression is also found in a subset of chronic lymphocytic leukemia patients with unmutated Ig genes and a poor clinical course [14]. LAT, a transmembrane adaptor protein expressed in T lymphocytes, natural killer cells and mast cells, is also an important mediator for TCR signaling [14]. Upon TCR engagement, activated ZAP-70 phosphorylates LAT at multiple conserved tyrosine residues within SH2 binding motifs, exposing these motifs as the docking sites for downstream signaling targets [14]. The phosphorylation process in LAT eventually leads to activation of the corresponding signaling pathways [14]. In regard to the positive staining for BAX, the BAX protein is a pro-apoptotic member of the Bcl-2 family of proteins; BAX is found in many cell types [15]. Bax is a cytosolic protein that translocates to the mitochondria and participates in Cytochrome c release in response to apoptotic stimuli [15]. Aberrant expression of BCL-2 family members can inappropriately promote or prevent apoptosis. Increased expression of BAX is associated with the apoptotic loss of neurons in Parkinson’s disease [15]. Given 1) the presence of junctional zone cytoid bodies in many cases of LE and 2) our BAX IHC findings, the role of BAX proapototic molecules in LE warrants further investigation. Based on our findings, the T cells and the cell signaling cascades are actively targeting not only the BMZ, but also skin adnexal structures; thus, the process may the result in dermal scarring and hair loss in some patients with LE. Additional studies are necessary to further explore and define these pathophysiologic possibilities.
REFERENCES
1. Obermoser G, Sontheimer RD, Zelger B: Overview of common, rare and atypical manifestations of cutaneous lupus erythematosus and histopathological correlates. Lupus. 2010; 19: 1050-1070. 2. Abreu-Velez AM, Howard MS, Hashimoto T, Grossniklaus HE: Human eyelid meibomian glands and tarsal muscle are recognized by autoantibodies from patients affected by a new variant of endemic pemphigus foliaceus in El-Bagre, Colombia, South America. J Am Acad Dermatol. 2010; 62: 437-447. 3. Abreu Velez AM, Howard MS, Hashimoto T: Palm tissue displaying a polyclonal autoimmune response in patients affected by a new variant of endemic pemphigus foliaceus in Colombia, South America. Eur J Dermatol. 2010; 20: 74- 71. 4. Howard MS, Yepes MM, Maldonado-Estrada JG, Villa- Robles E, Jaramillo A, Botero JH, et al: Broad histopathologic patterns of non-glabrous skin and glabrous skin from patients with a new variant of endemic pemphigus foliaceus-part 1. J Cutan Pathol. 2010; 37: 222- 230. 5. Abreu-Velez AM, Howard MS, Hashimoto K, Hashimoto T: Autoantibodies to sweat glands detected by different methods in serum and in tissue from patients affected by a new variant of endemic pemphigus foliaceus. Arch Dermatol Res. 2009; 301: 711-718. 6. Abreu Velez AM, Howard MS, Grossniklaus HE, Gao W, Hashimoto T: Neural system antigens are recognized by autoantibodies from patients affected by a new variant of endemic pemphigus foliaceus in Colombia. Clin Immunol. 2011; 31: 356-368. 7. Abreu-Velez AM, Klein AD, Howard MS: Junctional adhesion molecule overexpression in Kaposi varicelliform eruption skin lesions-as a possible herpes virus entry site. North Am J Med Sci 2010; 2: 433-437. 8. Abreu Velez AM, Smith JG Jr, Howard MS: Cutaneous lupus erythematosus with autoantibodies colocalizing with glial fibrillary acidic protein. N Dermatol Online. 2011; 2: 8-11. 9. Abreu Velez AM, Smith JG Jr, Howard MS: Vimentin compartamentalization in discoid lupus. North Am J Med Sci. 2010; 2: 106-110. 10. Abreu Velez AM, Girard JG, Howard MS: J. Antigen presenting cells in a patient with hair loss of and systemic lupus erythematosus. North Am J Med Sci. 2009; 1: 205- 210. 11. Albrecht J, Berlin JA, Braverman IM, Callen JP, Connolly MK, Costner MI, et al: Dermatology position paper on the revision of the 1982 ACR criteria for systemic lupus erythematosus. Lupus. 2004; 13: 839-849. 12. Hood AF, Farmer ER: Histopathology of cutaneous lupus erythematosus. Clinics in Dermatology. 1985; 3: 36- 38. 13. Xu L, Zhang L, Yi Y, Kang HK, Datta SK: Human lupus T cells resist inactivation and escape death by upregulating COX-2. Nature Medicine. 2004; 10: 411-415. 14. Paz PE, Wang S, Clarke H, Lu X, Stokoe D, Abo A; Mapping the Zap-70 phosphorylation sites on LAT (linker for activation of T cells) required for recruitment and activation of signaling proteins in T cells. Biochem J. 2001; 356: 461-471. 15. Narita M, Shimizu S, Ito T, Chittenden T, Lutz RJ, Matsuda H, et al: Bax interacts with the permeability transition pore to induce permeability transition and cytochrome c release in isolated mitochondria. Proc Natl Acad Sci USA 1998; 95: 14681-14686.
Comments are closed.